
15 joias da evolução


Um recurso da Nature para aqueles que queiram divulgar as evidências da evolução pela seleção natural.
Henry Gee, Rory Howlett e Phillip Campbell¹ Tradução de Eli Vieira²
A maioria dos biólogos aceitam como certa a ideia de que toda a vida evoluiu pela seleção natural ao longo de bilhões de anos. Eles pesquisam e ensinam em disciplinas que têm essa ideia como base, seguros de que a seleção natural é um fato, da mesma forma que é um fato que a Terra orbita o Sol. Dado que os conceitos e realidades da evolução de Darwin ainda estão sob ataque, embora raramente por biólogos, um resumo sucinto sobre o porquê de a evolução pela seleção natural ser um princípio empiricamente validado é útil para as pessoas terem à mão. Oferecemos aqui 15 exemplos publicados pela Nature ao longo da década passada para ilustrar a extensão, a profundidade e o poder do pensamento evolutivo. Nos alegramos de oferecer esse recurso gratuitamente e encorajamos sua livre divulgação.
1. Henry Gee é editor sênior da Nature, Rory Howlett é editor de consulta da Nature, Phillip Campbell é editor-chefe da Nature. 2. Eli Vieira é biólogo formado pela Universidade de Brasília e aluno de iniciação científica do Laboratório de Biologia Evolutiva, na mesma instituição. Este material é livre, conforme publicado na revista Nature em janeiro de 2009 (doi: 10.1038/nature07740).
JOIAS DO REGISTRO FÓSSIL 1. Ancestrais terrestres das baleias 2. Da água para a terra 3. A origem das penas 4. A história evolutiva dos dentes 5. A origem do esqueleto dos vertebrados
JOIAS DOS HABITATS 6. Seleção natural na especiação 7. Seleção natural em lagartos 8. Um caso de coevolução 9. Dispersão diferencial em aves selvagens 10. Sobrevivência seletiva em lebistes selvagens 11. A história evolutiva é importante
JOIAS DE PROCESSOS MOLECULARES 12. Os tentilhões das Galápagos e Darwin 13. Microevolução vai ao encontro da macroevolução 14. Resistência a toxinas em serpentes e em clame-da-areia 15. Variação versus estabilidade
JOIAS DO REGISTRO FÓSSIL
1 Ancestrais terrestres das baleias
Os fósseis oferecem pistas cruciais para a evolução, porque revelam as formas frequentemente notáveis de criaturas há muito desaparecidas na Terra. Alguns deles até documentam a evolução em ação, ao registrar criaturas em trânsito entre um ambiente e outro.
Baleias, por exemplo, estão belamente adaptadas à vida na água, e têm sido assim por milhões de anos. Mas como nós, são mamíferos. Elas respiram ar, dão à luz e amamentam seus filhotes. Mas há boas evidências de que os mamíferos evoluíram originalmente sobre a terra seca. Se é assim, então os ancestrais das baleias devem ter se mudado para a água em algum momento.
Acontece que temos fósseis numerosos dos aproximadamente dez milhões de anos iniciais da evolução das baleias. Nesses se incluem vários fósseis de criaturas aquáticas como Ambulocetus e Pakicetus, que têm características agora vistas apenas em baleias - especialmente a anatomia do ouvido interno - mas também têm membros como os dos mamíferos terrestres dos quais eles claramente derivaram. Tecnicamente, essas criaturas híbridas já eram baleias. O que estava faltando era o começo da história: as criaturas terrestres das quais as baleias evoluíram mais tarde.
Um trabalho publicado em 2007 pode ter apontado com precisão este grupo. Chamados de raoelídeos, essas criaturas agora extintas seriam parecidas com pequenos cachorrinhos, mas eram mais intimamente relacionadas aos ungulados com número par de dedos [artiodáctilos] - o grupo que inclui vacas, ovelhas, veados, porcos e hipopótamos modernos. Evidências moleculares também sugeriram que baleias e ungulados com número par de dedos compartilham uma profunda afinidade evolutiva.
O estudo detalhado de Hans Thewissen e colaboradores em NEOUCOM, Rootstown, mostra que um raoelídeo, Indohyus, é similar às baleias, mas diferente de outros artiodáctilos em relação à estrutura de seus ouvidos e dentes, à espessura de seus ossos e à composição química de seus dentes. Esses indicadores sugerem que essa criatura do tamanho de um guaxinim passava bastante tempo na água. Raoelídeos típicos, entretanto, têm uma dieta bem diferente da dieta das baleias, o que sugere que o estímulo para a mudança para a água pode ter sido a mudança de dieta.
Esse estudo demonstra a existência de potenciais formas transicionais no registro fóssil. Muitos outros exemplos poderiam ter sido ressaltados, e há toda razão para se pensar que muitos outros estão para serem descobertos, especialmente em grupos que estão bem representados no registro fóssil.
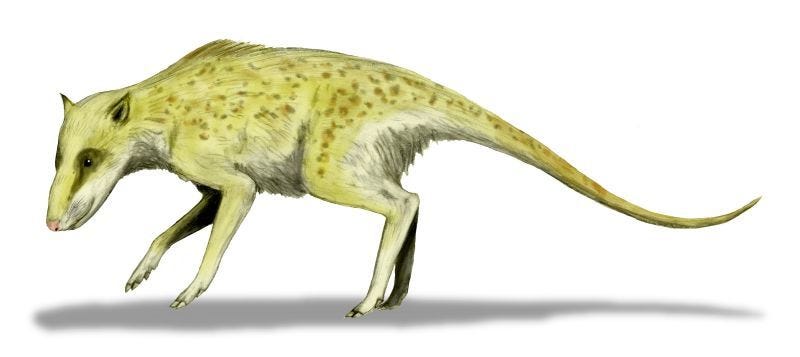
Indohyus major (CC 3.0 Arthur Weasley - Wikimedia Commons)
Referência Thewissen, J. G. M., Cooper, L. N., Clementz, M. T., Bajpai, S. & Tiwari, B. N. Nature 450, 1190–1194 (2007). Recursos adicionais Thewissen, J. G. M., Williams, E. M., Roe, L. J. & Hussain, S. T. Nature 413, 277–281 (2001). de Muizon, C. Nature 413, 259–260 (2001). Novacek, M. J. Nature 368, 807 (1994). Zimmer, C. At The Water’s Edge (Touchstone, 1999). Vídeo da pesquisa de Thewissen: www.nature.com/nature/videoarchive/ancientwhale Site do autor Hans Thewissen: www.neoucom.edu/DEPTS/ANAT/Thewissen
JOIAS DO REGISTRO FÓSSIL
2 Da água para a terra
De todos os animais, estamos mais familiarizados com os tetrápodos - eles são vertebrados (têm coluna vertebral) e vivem sobre a terra. Incluem-se nisso humanos, quase todos os animais domésticos e a maioria dos animais selvagens que uma criança reconheceria imediatamente: mamíferos, aves, anfíbios e répteis. A vasta maioria dos vertebrados, entretanto, não é de tetrápodos, mas de peixes. Há mais tipos de peixes, na verdade, que todas as espécies de tetrápodos somadas. De fato, pela lente da evolução, os tetrápodos são apenas mais um ramo da árvore genealógica dos peixes, um ramo cujos membros são adaptados para a vida fora da água.
A primeira transição da água para a terra aconteceu há mais de 360 milhões de anos. Foi uma das transições que necessitou de algumas das maiores mudanças já feitas na história da vida. Como nadadeiras se tornaram pernas? E como as criaturas transicionais lidaram com as formidáveis exigências da vida terrestre, da secura do ambiente à esmagadora gravidade?
Pensava-se que os primeiros terrícolas foram peixes que se encalhavam e evoluíram de modo a passar mais e mais tempo em terra firme, voltando à água para se reproduzirem. Ao longo dos últimos 20 anos, paleontólogos desenterraram fósseis que viraram essa ideia de cabeça para baixo. Os tetrápodos mais antigos, tais como o Acanthostega da Groenlândia oriental de cerca de 365 milhões de anos atrás, tinham pernas completamente formadas, com dedos, mas retiveram brânquias internas que se secariam facilmente se fossem expostas por muito tempo ao ar. Os peixes evoluíram pernas muito tempo antes de subirem à terra. Os tetrápodos mais antigos passaram a maior parte de sua evolução no ambiente aquático mais clemente. Subir à terra parece ter sido a última etapa.
Os pesquisadores suspeitam que os ancestrais dos tetrápodos eram criaturas chamadas elpistostegídeos. Esses peixes grandes, carnívoros de água rasa, se pareceriam bastante (até no comportamento) com jacarés ou salamandras gigantes. Eles aparentavam ser tetrápodos em muitos aspectos, exceto pelas nadadeiras que ainda tinham. Até recentemente, os elpistostegídeos eram conhecidos apenas por pequenos fragmentos de fósseis pobremente preservados, então foi difícil completar um esboço de como eles eram.
De poucos anos atrás até aqui, várias descobertas na ilha Ellesmere na região de Nunavut no norte do Canadá mudaram tudo isso. Em 2006, Edward Daeschler e seus colaboradores descreveram fósseis espetacularmente bem preservados de um elpistostegídeo conhecido como Tiktaalik, que nos permite reconstruir uma boa imagem de um predador aquático com similaridades distintas com os tetrápodos - de seu pescoço flexível à sua estrutura de barbatana semelhante a um membro.
A descoberta e a análise exaustiva do Tiktaalik iluminam a etapa anterior à evolução dos tetrápodos, e mostram como o registro fóssil é uma caixa de surpresas, embora as surpresas sejam completamente compatíveis com o pensamento evolutivo.

Tiktaalik roseae
Referências Daeschler, E. B., Shubin, N. H. & Jenkins, F A. Nature 440, 757–763 (2006). Shubin, N. H., Daeschler, E. B., & Jenkins, F A. Nature 440, 764–771 (2006). Recursos adicionais Ahlberg, P. E. & Clack, J. A. Nature 440, 747–749 (2006). Clack, J. Gaining Ground (Indiana Univ. Press, 2002) Shubin, N. Your Inner Fish (Allen Lane, 2008) Gee, H. Deep Time (Fourth Estate, 2000) Página de Tiktaalik: http://tiktaalik.uchicago.edu Sites dos autores Edward Daeschler: http://www.ansp.org/research/biodiv/vert_paleo/staff.php Neil Shubin: http://pondside.uchicago.edu/oba/faculty/shubin_n.html
JOIAS DO REGISTRO FÓSSIL
3 A origem das penas
Uma das objeções à teoria da evolução de Charles Darwin era a ausência de 'formas transicionais' no registro fóssil - formas que ilustrassem a evolução em ação, de um grande grupo de animais para outro. Entretanto, quase um ano após a publicação de Origem das Espécies, uma pena isolada foi achada no Jurássico Superior do calcário litográfico de Solnhofen (com idade de cerca de 150 milhões de anos), na Bavária. Em seguida, em 1861, foi encontrado o primeiro fóssil de Archaeopteryx, uma criatura com muitas características reptilianas primitivas, tais como dentes e uma longa cauda óssea, mas com asas e penas de voo, exatamente como uma ave.
Embora o Archaeopteryx fosse comumente visto como a primeira ave conhecida, muitos suspeitaram que ele fosse melhor descrito como um dinossauro com penas. Thomas Henry Huxley, colega e amigo de Darwin, discutiu a possível ligação evolutiva entre dinossauros e aves, e paleontólogos especularam, talvez com ousadia, que dinossauros com penas poderiam ser achados um dia.
Nos anos 80 [do século XX], depósitos do período do Cretáceo Inferior (cerca de 125 milhões de anos atrás) na província de Liaoning no norte da China sustentaram essas especulações da forma mais dramática, com descobertas de aves primitivas em abundância - junto com dinossauros com penas e com plumagem semelhante e penas. Começando com a descoberta do pequeno terópode Sinosauropteryx por Pei-ji Chen e colaboradores, do Instituto Nanjing de Geologia e Paleontologia da China, uma variedade de formas cobertas por penas foram encontradas em seguida. Muitos desses dinossauros com penas não podiam possivelmente voar, mostrando que as penas evoluíram primeiramente por outras razões que não o voo, possivelmente para displays sexuais [como faz o pavão] ou isolamento térmico, por exemplo. Em 2008, Fucheng Zhang e seus colegas da Academia Chinesa de Ciências em Pequim anunciaram a bizarra criatura Epidexipteryx, um pequeno dinossauro coberto por uma plumagem macia e dotado de quatro longas plumas em sua cauda. Os paleontólogos agora começam a pensar que suas especulações não eram ousadas o bastante, e que as penas eram na verdade bem comuns nos dinossauros.
A descoberta de dinossauros com penas não só apoiou a ideia das formas transicionais, como também mostrou que a evolução tem maneiras de fazer surgir uma fascinante variedade de soluções quando não fazíamos nem ideia de que havia problemas. O voo pode ter sido nada mais que uma oportunidade adicional que se apresentou a criaturas já revestidas por penas.

Archaeopteryx macrura (Vogt, C. Berlim, 1880)

Epidexipteryx sp. (CC 3.0 Arthur Weasley - Wikimedia Commons)
Referências Chen, P.-J., Dong, Z.-M. & Zhen, S.-N. Nature 391, 147–152 (1998). Zhang, F., Zhou, Z., Xu, X., Wang, X. & Sullivan, C. Nature 455, 1105–1008 (2008). Recursos adicionais Gee, H. (ed.) Rise of the Dragon (Univ. Chicago Press, 2001). Chiappe, L. Glorified dinosaurs (Wiley-Liss, 2007). Gee, H. & Rey, L. V. A Field Guide to Dinosaurs (Barron’s Educational, 2003).
JOIAS DO REGISTRO FÓSSIL
4 A história evolutiva dos dentes
Uma motivação para o estudo do desenvolvimento é a descoberta de mecanismos que guiam a mudança evolutiva. Kathryn Kavanagh e seus colaboradores, da Universidade de Helsinki, investigaram justamente isso observando os mecanismos por trás do tamanho relativo e número de dentes molares em camundongos. A pesquisa, publicada em 2007, desvendou o padrão de expressão dos genes que governam o desenvolvimento dos dentes - os molares emergem da parte da frente para a parte de trás, com cada novo dente sendo menor que o último a emergir.
A beleza deste estudo está em sua aplicação. O modelo dos pesquisadores prevê os padrões de dentição encontrados em espécies de roedores semelhantes aos camundongos, com várias dietas, fornecendo um exemplo de evolução ecologicamente guiada por uma trajetória favorecida no desenvolvimento. Em geral, o trabalho mostra como o padrão de expressão gênica pode ser modificado durante a evolução para produzir mudanças adaptativas em sistemas naturais.
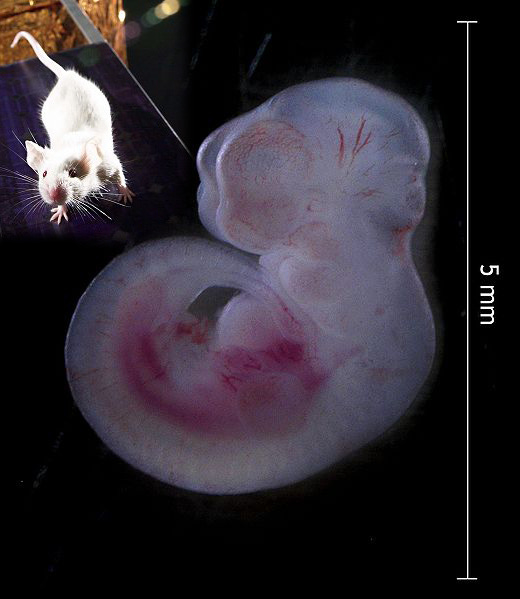
Um adulto e um embrião de 12,5 dias de Mus musculus (camundongo).
Referência Kavanagh, K. D., Evans, A. R. & Jernvall, J. Nature 449, 427–432 (2007). Recursos adicionais Polly, P. D. Nature 449, 413–415 (2007). Evans, A. R., Wilson, G. P., Fortelius, M. & Jernvall, J. Nature 445, 78–81 (2006). Kangas, A. T., Evans, A. R., Thesleff, I. & Jernvall, J. Nature 432, 211–214 (2004). Jernvall, J. & Fortelius, M. Nature 417, 538–540 (2002). Theodor, J. M. Nature 417, 498–499 (2002). Site do autor Jukka Jernvall: http://www.biocenter.helsinki.fi/bi/evodevo
JOIAS DO REGISTRO FÓSSIL
5 A origem do esqueleto dos vertebrados
Devemos muito do que nos faz humanos a um notável tecido chamado crista neural, encontrado apenas em embriões. As células da crista neural emergem da medula espinhal prematura e migram para todo o corpo, efetuando uma série de transformações importantes. Sem a crista neural, não teríamos a maioria dos ossos em nossa face e pescoço, e muitas das características de nossa pele e de nossos órgãos sensoriais. A crista neural parece ser exclusiva dos vertebrados, e ajuda a explicar por que os vertebrados têm 'cabeças' e 'faces' distintas.
Desvendar a história evolutiva da crista neural é especialmente difícil em formas fósseis, pois dados embrionários estão obviamente ausentes. Um mistério crucial, por exemplo, é saber quanto do crânio dos vertebrados provém da contribuição das células da crista neural e quanto vem de camadas mais internas de tecido.
Novas técnicas permitiram aos pesquisadores rotular e acompanhar células individuais enquanto os embriões se desenvolviam. As técnicas revelaram, até o nível das células individuais, que as bordas dos ossos do pescoço e do ombro derivam da crista neural. O tecido derivado da crista neural ancora a cabeça na superfície anterior da cintura escapular, enquanto o esqueleto que forma a parte de trás do pescoço e dos ombros [escápulas e clavículas] cresce de uma camada mais interna de tecido chamada mesoderme.
Tal mapeamento detalhado, em animais vivos, esclarece a evolução de estruturas em cabeças e pescoços de animais extintos há tempos, mesmo sem tecido mole fossilizado, como pele e músculo. Similaridades de esqueleto que resultam de uma história evolutiva compartilhada podem ser identificadas nas marcas de inserção musculares [nos ossos]. Isso permite delinear, por exemplo, a localização do maior osso do "ombro" de ancestrais terrestres extintos dos vertebrados, o cleitro. Esse osso parece sobreviver até hoje na forma de parte da escápula em mamíferos viventes.
Esse tipo de exame evolutivo pode ter relevância clínica imediata. As partes do esqueleto identificadas como derivadas da crista neural, por Toshiyuki Matsuoka e seus colaboradores no Instituto Wolfson para Pesquisa Biomédica em Londres, são especificamente afetadas em várias doenças do desenvolvimento em humanos, o que permite esclarecimentos sobre suas origens.
O estudo de Matsuoka mostra como uma análise detalhada da morfologia dos animais viventes, informada pelo pensamento evolutivo, ajuda os pesquisadores a interpretar formas fossilizadas e extintas.

Cintura escapular humana, vista dorsal (Gray's Anatomy Of The Human Body, 1918)
Referência Matsuoka, T. et al. Nature 436, 347–355 (2005). Site do autor Georgy Koentges: http://www2.warwick.ac.uk/fac/sci/systemsbiology
JOIAS DOS HABITATS
6 Seleção natural na especiação
A teoria evolutiva prevê que a seleção natural disruptiva terá frequentemente um papel principal na especiação. Trabalhando com esgana-gatas (Gasterosteus aculeatus [peixes da família Gasterosteidae]), Jeffrey McKinnon e seus colaboradores, na Universidade de Wisconsin em Whitewater, relataram em 2004 que o isolamento reprodutivo pode evoluir como um subproduto da seleção sobre o tamanho do corpo. Esse trabalho fornece um vínculo entre o estabelecimento do isolamento reprodutivo e a divergência de uma característica ecologicamente importante.
O estudo foi feito numa escala geográfica extraordinária, envolvendo testes de acasalamento entre peixes coletados no Alasca, Colúmbia Britânica, Islândia, Reino Unido, Noruega e Japão; e se sustentou em análises de genética molecular que forneceram sólidas evidências de que os peixes que se adaptaram a viver em cursos d'água evoluíram repetidamente de ancestrais marinhos, ou de peixes que vivem no oceano mas retornam à água doce para reprodução. Tais populações migratórias no estudo tinham em média corpos maiores se comparadas às que vivem em cursos d'água. Os indivíduos tenderam a se acasalar com peixes de tamanho similar ao seu, o que serve bem para o isolamento reprodutivo entre diferentes ecótipos [populações geneticamente singulares adaptadas a seu ambiente local] de cursos d'água e seus vizinhos próximos de vida marítima.
Levando em consideração as relações evolutivas, uma comparação dos vários tipos de esgana-gata, quer de cursos d'água ou marinhos, apóia fortemente a visão de que a adaptação a diferentes ambientes acarreta o isolamento reprodutivo. Os experimentos dos pesquisadores confirmaram também a conexão entre divergência de tamanho e o estabelecimento de isolamento reprodutivo - embora outras características além do tamanho também contribuam para o isolamento reprodutivo em alguma medida.

Esgana-gata (Gasterosteus aculeatus)
Referência McKinnon, J. S. et al. Nature 429, 294–298 (2004). Recursos Adicionais Gillespie, R. G. & Emerson, B. C. Nature 446, 386–387 (2007). Kocher, T. D. Nature 435, 29–30 (2005). Emerson, B. C. & Kolm, N. Nature 434, 1015–1017 (2005). Sites dos autores Jeffrey McKinnon: http://facstaff.uww.edu/mckinnoj/mckinnon.html David Kingsley: http://kingsley.stanford.edu Dolph Schluter: http://www.zoology.ubc.ca/~schluter
JOIAS DOS HABITATS
7 Seleção natural em lagartos
Uma hipótese evolutiva popular diz que mudanças comportamentais em novos ambientes anulam os efeitos da seleção natural. Mas o trabalho de Jonathan Losos e seus colaboradores na Universidade de Harvard, em 2003, empresta pouco apoio a essa teoria. Os pesquisadores introduziram um grande lagarto terrícola e predador, Leiocephalus carinatus, a seis ilhas pequenas nas Bahamas, com seis outras ilhas servindo como controle. Descobriram que a presa desse lagarto, que é um lagarto menor chamado Anolis sagrei, passava mais tempo em maiores alturas na vegetação de ilhas ocupadas pelo predador do que em ilhas onde L. carinatus estava ausente. Mas a mortalidade de A. sagrei continuou mais alta nas ilhas experimentais que nas ilhas controle.
A presença do predador maior selecionou em favor de machos de lagarto A. sagrei com pernas mais altas, que podem correr mais rápido, e também favoreceu fêmeas maiores, que são tanto mais rápidas quanto mais difíceis de vencer e de ingerir. Os pesquisadores não detectaram seleção de tamanho em machos; sugeriram que os machos maiores podem ter sido mais vulneráveis por causa de seu comportamento territorial conspícuo.
O estudo mostra como a introdução de um predador pode fazer com que os indivíduos de uma espécie predada mudem seu comportamento de modo a reduzir o risco de predação, mas também causa uma resposta evolutiva no nível da população, resposta que difere entre os sexos de acordo com sua ecologia.

Leiocephalus carinatus (CC 3.0 Ianaré Sévi - Wikimedia Commons) Anolis sagrei
Referência Losos, J. B., Schoener, T. W. & Spiller, D. A. Nature 432, 505–508 (2004). Recursos adicionais Butler, M. A., Sawyer, S. A. & Losos, J. B. Nature 447, 202–205 (2007). Kolbe, J. J. et al. Nature 431, 177–181 (2004). Calsbeek, R. & Smith, T. B. Nature 426, 552–555 (2003). Losos, J. B. et al. Nature 424, 542–545 (2003). Site do autor Jonathan Losos: http://www.oeb.harvard.edu/faculty/losos/jblosos
JOIAS DOS HABITATS
8 Um caso de coevolução
As espécies evoluem juntas, e em competição. Predadores evoluem sempre armas e habilidades mais letais para caçarem suas presas, que por sua vez, como resultado da canônica 'luta pela existência' de Darwin, se tornam melhores em escapar, e então a corrida armamentista continua. Em 1973, o biólogo evolutivo Leigh Van Valen comparou essa corrida ao comentário da Rainha Vermelha para Alice em "Alice no País do Espelho", de Lewis Carroll: "é preciso correr o máximo que pode, para continuar no mesmo lugar. Se você quer chegar a outro lugar, precisa correr pelo menos duas vezes mais rápido que isso!" Nascia a hipótese da 'Rainha Vermelha', sobre a coevolução.
Um problema em estudar a dinâmica de Rainha Vermelha é que essa dinâmica só pode ser vista no eterno presente. Descobrir sua história é problemático, porque a evolução geralmente eliminou todos os estágios anteriores.
Felizmente, Ellen Decaestecker e seus colegas, na Universidade Católica de Leuven na Bélgica, descobriram uma notável exceção na corrida armamentista coevolutiva entre as pulgas d'água (Daphnia) e os parasitas microscópicos que as infestam; a pesquisa foi publicada em 2007. Enquanto as pulgas d'água se tornam melhores em evitar o parasitismo, os parasitas se tornam melhores em infectá-las. Ambos presa e predador, nesse sistema, podem persistir em estágios dormentes por muitos anos na lama do fundo do lago que compartilham. Os sedimentos do lago podem ser datados conforme o ano em que se formaram, e os predadores e presas enterrados podem ser ressuscitados. Assim, suas interações podem ser testadas, uma contra a outra, e contra predadores e presas provenientes de seus passados e futuros relativos.
Confirmando as expectativas teóricas, o parasita adaptou-se a seu hospedeiro ao longo de apenas alguns anos. Sua infectividade a qualquer determinado tempo mudou pouco, mas sua virulência e aptidão [fitness] cresceram constantemente - equiparadas em cada etapa pela capacidade das pulgas d'água de resistência a elas.
Esse estudo fornece um exemplo elegante no qual um registro histórico de alta resolução do processo coevolutivo providenciou uma confirmação da teoria evolutiva, mostrando que a interação entre parasitas e seus hospedeiros não é fixa no tempo, mas, alternativamente, é o resultado de uma corrida armamentista dinâmica de adaptação e contra-adaptação, conduzida pela seleção natural de geração para geração.
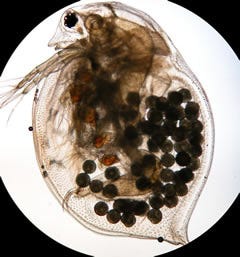
Daphnia magna
Referência Decaestecker, E. et al. Nature 450, 870–873 (2007). Recursos adicionais The Red Queen Hypothesis: http://en.wikipedia.org/wiki/Red_Queen Van Valen, L. Evol. Theory 1, 1–30 (1973). Site do autor Ellen Decaestecker: http://bio.kuleuven.be/de/dea/people_detail.php?pass_id=u0003403
JOIAS DOS HABITATS
9 Dispersão diferencial em aves selvagens
O fluxo gênico causado pela migração, por exemplo, pode desfazer adaptações para condições locais e ir na contramão da diferenciação evolutiva dentro de uma população e entre populações. De fato, a teoria clássica da genética de populações sugere que quanto mais as populações locais migrarem e se intercruzarem, mais geneticamente similares elas serão. Esse conceito parece estar de acordo com o senso comum, assume que o fluxo gênico é um processo casual, como a difusão. Mas a dispersão não-casual pode na verdade favorecer a adaptação local e a diferenciação evolutiva, como relataram em 2005 Ben Sheldon e seus colaboradores, do Instituto Edward Gray de Ornitologia de Campo, em Oxford, Reino Unido.
O trabalho deles foi parte de um estudo multidécada sobre os chapins-reais (Parus major) que habitam um bosque em Oxfordshire, Reino Unido. Os pesquisadores descobriram que a quantidade e tipo de variação genética no peso dos ninhegos dessa ave canora difere entre uma parte do bosque e outra. Esse padrão de variação leva a respostas diversas à seleção em partes diferentes do bosque, resultando em adaptação local. O efeito é reforçado pela dispersão não-casual; aves individuais selecionam e se reproduzem em diferentes habitats de um modo que aumenta sua aptidão [fitness]. Os autores concluem que "quando o fluxo gênico não é homogêneo, a diferenciação evolutiva pode ser rápida e pode ocorrer sobre escalas espaciais surpreendentemente pequenas."
Em outro estudo sobre chapins-reais na ilha Vlieland da Holanda, publicado na mesma edição da Nature, Erik Postma e Arie van Noordwijk, do Instituto Holandês de Ecologia em Heteren, descobriram que o fluxo gênico mediado pela dispersão não-casual mantém uma grande diferença genética do tamanho das garras numa pequena escala espacial, mais uma vez ilustrando, como dizem esses cientistas, "o grande efeito da imigração na evolução de adaptações locais e na estrutura genética da população".

Chapim-real fêmea (Parus major - Martin Olsson, Wikimedia Commons)
Referências Garant, D., Kruuk, L. E. B., Wilkin, T. A., McCleery, R. H. & Sheldon, B. C. Nature 433, 60–65 (2005). Postma, E. & van Noordwijk, A. J. Nature 433, 65-68 (2005). Recursos adicionais Coltman, D. W. Nature 433, 23–24 (2005). Sites dos autores Ben Sheldon: http://www.zoo.ox.ac.uk/egi/people/faculty/ben_sheldon.htm Erik Postma: http://www.nioo.knaw.nl/ppages/epostma Arie van Noordwijk: http://www.nioo.knaw.nl/PPAGES/avannoordwijk David Coltman: http://www.biology.ualberta.ca/faculty/david_coltman
JOIAS DOS HABITATS
10 Sobrevivência seletiva em lebistes selvagens
A seleção natural favorece características que aumentam a aptidão. Ao longo do tempo, poderia-se esperar que tal seleção exaurisse a variação genética por conduzir variantes genéticas vantajosas à fixação em detrimento de variantes menos vantajosas ou deletérias. Na verdade, as populações naturais apresentam frequentemente grandes quantidades de variação genética. Então como esta é mantida?
Um exemplo é o polimorfismo genético que se vê nos padrões de cor de machos de lebiste (Poecilia reticulata [peixe da família Poeciliidae]). Como relatado em 2006, Kimberly Hughes e colaboradores, da Universidade de Illinois em Urbana-Champaign, manipularam as frequências de machos com diferentes padrões de cor em três populações selvagens de lebiste em Trinidad. Os pesquisadores mostraram que variantes raras têm taxas muito maiores de sobrevivência do que os mais comuns. Essencialmente, variantes são favorecidas quando raras, e selecionadas negativamente quando comuns.
Tal sobrevivência 'dependente de frequência', na qual a seleção favorece tipos raros, tem sido relacionada à manutenção de polimorfismos moleculares, morfológicos e relativos à saúde em humanos e em outros mamíferos.

Lebiste macho (Poecilia reticulata)
Referência Olendorf, R. et al. Nature 441, 633–636 (2006). Recurso adicional Foerster, K. et al. Nature 447, 1107–1110 (2007). Sites dos autores Kimberly Hughes: http://www.bio.fsu.edu/faculty-hughes.php Anne Houde: http://www.lakeforest.edu/academics/faculty/houde David Reznick: http://www.biology.ucr.edu/people/faculty/Reznick.html
JOIAS DOS HABITATS
11 A história evolutiva é importante
Pensa-se, muitas vezes, que a evolução tem algo a ver com encontrar soluções ótimas para os problemas que a vida apresenta. Mas a seleção natural só pode trabalhar com os materiais disponíveis - materiais que são por sua vez os resultados de muitos milhões de anos de história evolutiva. Ela nunca começa com uma folha em branco. Se o ótimo fosse o caso, então os tetrápodos, ao se depararem com o problema de andar sobre a terra, poderiam talvez ter evoluído rodas em vez de terem suas nadadeiras transformadas em pernas.
Um caso real da engenhosidade da adaptação diz respeito a uma moreia (Muraena retifera), um predador dos recifes de coral que parece uma grande serpente. Historicamente, os peixes ósseos usam a sucção para capturar suas presas. Um peixe, ao se aproximar do alimento, abre sua boca para criar uma grande cavidade que suga água para dentro de si junto com a presa. Quando o excesso de água sai pelas fendas branquiais, o peixe leva a presa ao esôfago e às mandíbulas faringeais, que são um segundo conjunto de mandíbulas e dentes derivados do esqueleto que sustenta as brânquias. Mas as moreias têm um problema por causa de sua forma alongada e estreita. Mesmo com suas mandíbulas abertas, sua cavidade bucal é pequena demais para gerar sucção suficiente para conduzir a presa até suas mandíbulas faringeais. A solução para este impasse foi documentada em 2007.
Através de observação cuidadosa e de cinematografia de raios-X, Rita Mehta e Peter Wainwright da Universidade da Califórnia, Davis, descobriram a empolgante solução da evolução. Em vez de a presa vir às mandíbulas faringeais, as mandíbulas faringeais se projetam para a cavidade bucal, encurralando a presa e arrastando-a para dentro. Esse, dizem os pesquisadores, é o primeiro caso descrito de um vertebrado usando um segundo conjunto de mandíbulas para conter e transportar a presa, e é a única alternativa conhecida para o transporte hidráulico de presa registrado na maioria dos peixes ósseos - uma grande inovação que pode ter contribuído para o sucesso das moreias como predadores.
A mecânica das mandíbulas faringeais da moréia lembra os mecanismos de retenção usados pelas serpentes - também criaturas longas, estreitas, que fazem predação. Esse é um exemplo da convergência, o fenômeno evolutivo no qual criaturas distantemente aparentadas evoluem soluções similares para problemas comuns.
Esse estudo demonstra a natureza contingente da evolução; como um processo ela não pode desfrutar do luxo de 'projetar do nada'.

Moreia
Referência Mehta, R. S. & Wainwright, P. C. Nature 449, 79–82 (2007). Recurso adicional Westneat, M. W. Nature 449, 33–34 (2007). Sites dos autores Rita Mehta: http://www.eve.ucdavis.edu/~wainwrightlab/rsmehta/index.html Peter Wainwright: http://www.eve.ucdavis.edu/~wainwrightlab
JOIAS DE PROCESSOS MOLECULARES
12 Os tentilhões das Galápagos e Darwin
Quando Charles Darwin visitou as Ilhas Galápagos, registrou a presença de várias espécies de tentilhão que tinham aparência muito similar, exceto por seus bicos. Tentilhões do solo têm bicos profundos e largos; tentilhões do cacto têm bicos longos e pontudos; tentilhões-rouxinóis têm bicos afilados e pontudos; refletindo diferenças em suas respectivas dietas. Darwin especulou que todos os tentilhões tinham um ancestral comum que tinha migrado para as ilhas. Parentes próximos dos tentilhões das Galápagos são conhecidos no continente da América do Sul, e o caso dos tentilhões de Darwin se tornou desde então o exemplo clássico de como a seleção natural levou à evolução de uma variedade de formas adaptadas a nichos ecológicos diferentes a partir de uma espécie ancestral comum - o que se chama de 'radiação adaptativa'. Essa ideia tem sido fortalecida desde então, por dados que mostram que mesmo pequenas diferenças na profundidade, largura e comprimento do bico podem ter grandes consequências para a aptidão geral das aves.
Para descobrir que mecanismos genéticos estão na base das mudanças no formato do bico que marca cada espécie, Arhat Abzhanov, da Universidade de Harvard, e seus colaboradores examinaram genes numerosos que são ativados durante o desenvolvimento dos bicos nos filhotes de tentilhão; o estudo foi publicado em 2006. Os pesquisadores descobriram que as diferenças na forma coincidem com a expressão diferenciada do gene para calmodulina, uma molécula envolvida na sinalização por cálcio, que é vital em muitos aspectos do desenvolvimento e do metabolismo. A calmodulina é expressada mais fortemente nos bicos longos e pontudos dos tentilhões do cacto do que em bicos mais robustos de outras espécies. Aumentar artificialmente a expressão da calmodulina nos tecidos embrionários que dão origem ao bico causa um alongamento da parte superior do bico, parecido com o que se observa em tentilhões do cacto. Os resultados mostram que ao menos parte da variação na forma do bico dos tentilhões de Darwin é provável que esteja relacionada à variação na atividade da calmodulina, e associam a calmodulina ao desenvolvimento de estruturas craniofaciais do esqueleto de maneira mais geral.
O estudo mostra como os biólogos estão indo além da mera documentação de mudanças evolutivas para identificar os mecanismos moleculares subjacentes a elas.

Tentilhões das galápagos (por John Gould)
Referência Abzhanov, A. et al. Nature 442, 563–567 (2006). Sites dos autores Clifford Tabin: http://www.hms.harvard.edu/dms/bbs/fac/tabin.html Peter Grant: http://www.eeb.princeton.edu/FACULTY/Grant_P/grantPeter.html
JOIAS DE PROCESSOS MOLECULARES
13 Microevolução vai ao encontro da macroevolução
Darwin concebeu a mudança evolutiva como um acontecimento dividido em passos pequenos, infinitesimais. Ele os chamou de 'gradações imperceptíveis' que, se extrapoladas a longos períodos de tempo, resultariam em mudanças radicais de forma e função. Há uma montanha de evidências para essas pequenas mudanças, chamadas de microevolução - a evolução da resistência a antibióticos, por exemplo, é apenas um de muitos casos documentados.
Podemos inferir do registro fóssil que mudanças maiores de espécie para espécie, ou macroevolução, também ocorrem, mas são naturalmente mais difíceis de se observar em ação. Porém, os mecanismos da macroevolução podem ser vistos no aqui e agora, na arquitetura dos genes. Às vezes os genes envolvidos nas vidas cotidianas dos organismos estão conectados ou são os mesmos que governam características principais na forma e no desenvolvimento dos animais. Então a evolução mais vulgar pode ter efeitos extensos.
Sean Carroll, do Instituto Médico Howard Hughes em Chevy Chase, Maryland, e seus colaboradores observaram um mecanismo molecular que contribui para o ganho de uma única mancha nas asas de machos da mosca da espécie Drosophila biarmipes; eles relataram seus achados em 2005. Os pesquisadores mostraram que a evolução dessa mancha é conectada a modificações de um elemento regulatório ancestral de um gene envolvido na pigmentação. Esse elemento regulatório adquiriu, com o tempo, sítios de ligação para fatores de transcrição que são componentes primitivos do desenvolvimento das asas. Um dos fatores de transcrição que se liga especificamente ao elemento regulatório do gene yellow é codificado pelo gene engrailed, que é um gene fundamental para o desenvolvimento como um todo.
Isso mostra que um gene envolvido em um processo pode ser cooptado para outro, em princípio conduzindo a mudança macroevolutiva.
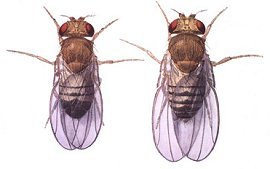
Drosophila (macho e fêmea)
Referência Gompel, N., Prud’homme, B., Wittkopp, P. J., Kassner, V. A. & Carroll, S. B. Nature 433, 481–487 (2005). Recursos adicionais Hendry, A. P. Nature 451, 779–780 (2008). Prud’homme, B. et al. Nature 440, 1050–1053 (2006). Site do autor Sean Carroll: http://www.hhmi.org/research/investigators/carroll_bio.html
JOIAS DE PROCESSOS MOLECULARES
14 Resistência a toxinas em serpentes e em clame-da-areia
Os biólogos estão compreendendo cada vez mais os mecanismos moleculares subjacentes à mudança evolutiva adaptativa. Em algumas populações do tritão Taricha granulosa, por exemplo, os indivíduos acumulam o veneno neuroativo tetrodotoxina em sua pele, aparentemente como uma defesa contra a cobra Thamnophis sirtalis. As cobras dessa espécie que predam os tritões venenosos evoluíram resistência à toxina. Através de trabalho exaustivo, Shana Geffeney, da Escola de Medicina de Stanford, na Califórnia, e seus colaboradores descobriram o mecanismo por trás disso; seu estudo foi publicado em 2005. A variação no nível de resistência das cobras à sua caça venenosa pode ser delineada em mudanças moleculares que afetam a ligação da tetrodotoxina a um canal de sódio particular.
Uma seleção similar para a resistência a toxinas aparentemente ocorre nos clames-da-areia (Mya arenaria) em áreas costeiras do Atlântico Norte no continente americano, como relatado por Monica Bricelj do Instituto de Biociências Marinhas da Nova Escócia, Canadá, e seus colaboradores na mesma edição da Nature. As algas que produzem 'marés vermelhas' geram saxitoxina - uma causa de envenenamento por frutos-do-mar em humanos. Esses molucos se expõem à toxina ao ingerirem as algas. Os provenientes de áreas com marés vermelhas recorrentes são relativamente resistentes à toxina e acumulam-na em seus tecidos. Os que vivem em áreas não afetadas não evoluíram tal resistência.
A resistência à toxina nas populações expostas está correlacionada com uma única mutação no gene que codifica para um canal de sódio, num sítio já relacionado à ligação da saxitoxina. Parece provável, portanto, que a saxitoxina age como um potente agente seletivo nos clames-da-areia e leva à adaptação genética.
Esses dois estudos mostram como pressões seletivas similares podem levar a respostas adaptativas similares mesmo em táxons [unidades de classificação] muito diferentes.

Taricha granulosa

Thamnophis sirtalis (CC 2.0 Steve Jurvetson - Wikimedia Commons)

Mya arenaria
Referências Geffeney, S. L., Fujimoto, E., Brodie, E. D., Brodie, E. D. Jr, & Ruben, P. C. Nature 434, 759–763 ( 2005). Bricelj, V. M. et al. Nature 434, 763–767 (2005). Recursos adicionais Mitchell-Olds, T. & Schmitt, J. Nature 441, 947–952 (2006). Bradshaw, H. D. & Schemske, D. W. Nature 426, 176–178 (2003). Coltman, D. W., O’Donoghue, P, Jorgenson, J. T., Hogg, J. T. Strobeck, C. & Festa-Bianchet, M. Nature 426, 655–658 (2003). Harper Jr, G. R. & Pfennig, D. W. Nature 451, 1103–1106 (2008). Ellegren, H. & Sheldon, B. Nature 452, 169–175 (2008). Sites dos autores Shana Geffeney: http://wormsense.stanford.edu/people.html Monica Bricelj: http://marine.biology.dal.ca/Faculty_Members/Bricelj,_Monica.php
JOIAS DE PROCESSOS MOLECULARES
15 Variação versus estabilidade
As espécies podem permanecer sem grandes mudanças por milhões de anos, tempo o bastante para coletarmos suas marcas no registro fóssil. Mas elas mudam também, e muitas vezes muito abruptamente. Isso levou alguns a se perguntarem se as espécies - geralmente aquelas se desenvolvendo em trajetórias específicas - guardavam oculto o potencial para a mudança abrupta, soltando uma enxurrada de variação em tempos de estresse ambiental, variação esta sobre a qual a seleção pode atuar, e que fica encoberta em outras circunstâncias em que não há estresse.
Essa ideia de 'capacidade evolutiva' foi trazida à baila primeiramente por Suzanne Rutherford e Susan Lindquist em experimentos surpreendentes com moscas-das-frutas. Sua ideia era que proteínas-chave envolvidas na regulação de processos de desenvolvimento são 'acompanhadas' por uma proteína chamada Hsp90, que é produzida mais em tempos de estresse. Nessas situações, Hsp90 é sobrepujada por outros processos e as proteínas normalmente reguladas por ela podem circular livremente, produzindo um rebuliço de variação que de outra forma estaria oculta.
Aviv Bergman, da Faculdade Albert Einstein de Medicina em Nova York, e Mark Siegal, da Universidade de Nova York, exploraram se a capacidade evolutiva é restrita à Hsp90 ou é encontrada mais geralmente; seu estudo foi publicado em 2003. Eles usaram simulações numéricas de redes genéticas complexas e dados de expressão gênica de todo o genoma de linhagens de levedura nas quais genes individuais foram deletados. Mostraram que a maioria e talvez todos os genes guardem variação em reserva que é liberada apenas quando são funcionalmente comprometidos. Em outras palavras, parece que a capacidade evolutiva pode ser maior e mais profunda do que se vê na Hsp90.
Modelo de dímero da proteína Hsp90 (heat shock protein 90, ou proteína de choque térmico 90)
Referências Bergman, A. & Siegal, M. L. Nature 424, 549–552 (2003). Recursos adicionais Stearns, S. C. Nature 424, 501–504 (2003). Rutherford, S. L. & Lindquist, S. Nature 396, 336–342 (1998). Sites dos autores Mark Siegal: http://www.nyu.edu/fas/biology/faculty/siegal/index.html Aviv Bergman: http://www.bergmanlab.org Susan Lindquist: http://www.wi.mit.edu/research/faculty/lindquist.html Suzanne Rutherford: http://depts.washington.edu/mcb/facultyinfo.php?id=142 Stephen Stearns: http://www.yale.edu/eeb/stearns
Para mais informações sobre evolução e Darwin, visite: www.nature.com/darwin (em inglês)
http://evolucionismo.ning.com (em português)
O tradutor agradece pelo apoio das seguintes instituições:

Universidade de Brasília Instituto de Ciências Biológicas
Departamento de Genética e Morfologia
Laboratório de Biologia Evolutiva
Brasília, janeiro de 2009.
www.nature.com/evolutiongems
evolucionismo.org